
You are using an out of date browser. It may not display this or other websites correctly.
You should upgrade or use an alternative browser.
You should upgrade or use an alternative browser.
Doubled haploids
- Thread starter mofeta
- Start date
...meiosis being the creation of the haploid cells.
Now I see your point, sorry I didn't understand what you meant. I thought you meant in the haploid plant's progeny.
You are correct, of course. I misspoke earlier. I shouldn't have said "avoid the reshuffling of meiosis". The haploid cells are created through this reshuffling/meiosis. What I meant is that the doubling of the haploid cells avoids any reshuffling of the genome of the haploid.
In meiosis a lot more than reshuffling takes place. It's when the rewrites take place. The doubling of segments of dna, the deletion of segments, and the random swaps and passing down the line of whole genes. And all randomly, and different in each haploid cell produced. ~snip~ the haploid cells produced in any plant, will not only be diff from the original, but also from each other. ~snip~ But you still aren't getting seeds that contain the exact same instruction sets or the same order that existed in the mother DH plant.
Basically correct.
The point is that some of the haploid gametes (maybe very few) will produce individuals that are phenotypically very similar to the desirable parent. When doubled, they are then perfectly homozygous for the desirable trait(s).
diploid seeds created which inherit (and I do accept the concept but still) diploid not DiHaploid DNA strands, one from original DH mom and one from pollen producing mom. And the process is the same in producing the ovums you pollinate, so each strand of dna is now different from the original DH mom's. And these will most certainly never be identical changes in both the pollen and the ovums that meet to produce the seeds.
This is also basically correct. DH is not a way of identically "copying" the desirable parent. It is a way of perfectly copying the genome of a selected (probably from many many) haploid individual into a perfectly homozygous diploid plant.
I'm sorry, but I don't have time to discuss anymore right now. I will write more when I can. You obviously have a good grasp of this stuff and I look forward to more discussion.
Hey a quick note-
Maybe our discussion can go beyond DH as a breeding technique and into molecular genetics in general.
I'd like to discuss the relation between the concepts of a strand of DNA, a double helix of DNA, a coding strand, a template strand, a chromosome, and ploidy of a cell.
Maybe our discussion can go beyond DH as a breeding technique and into molecular genetics in general.
I'd like to discuss the relation between the concepts of a strand of DNA, a double helix of DNA, a coding strand, a template strand, a chromosome, and ploidy of a cell.
Keep on chatting people, this is rad to read ") ! Sorry I don't have anything to add and I have no knowledge in this field but, thanks, I'm enjoying all this information!
! Sorry I don't have anything to add and I have no knowledge in this field but, thanks, I'm enjoying all this information!
! Sorry I don't have anything to add and I have no knowledge in this field but, thanks, I'm enjoying all this information!I really can't believe how annoying this site is, I typed up a massive response and lost it when I was told I was no longer logged into the site. That shit really makes posting here so frustrating and now I have to start all over again.
Hey Mofeta, its really nice to be able to discuss this stuff again, so many who used to hold these conversations have now gone awol from this site. And ignoring the politics side of things, just the mechanical difficulties I just ran into makes it so frustrating at times. Rewriting long technical posts is really annoying.
Mofeta : "What I meant is that the doubling of the haploid cells avoids any reshuffling of the genome of the haploid." yes I do agree that the adding of poison (providing you get the dosage just right for the Haploid plant) will merely create a DiHaploid and not a diploid.
Damn I can't remember what I wrote now lol.
The problem as I see it, is that most of the traits in the canna world, aren't necessarily dominant or recessive, but rather co-dominant. And when working with only one side of the double helix, you are automatically excluding the possibility of those traits being present in the DiHaploid. This also ignores those traits that rely on more than one co-dominant gene, of which I believe there are many. Accepting that (to use your original example) the GS Cookies plant, is not a DiHaploid plant, but rather a diploid, then to try to replicate that plant using haploid cells will exclude half of the dna instruction sets necessary to create the plant.
When you say basically correct, if there is anything at all that you disagree with, please just say so straight away, forget about ego issues, I'm not gonna let that get in the way of a decent discussion at all. And it's necessary to get diffs identified instantly in a discussion of this type as letting them slide will only amplify difficulties later on in the discussion.
Sorry man, I just can't remember what I wrote at all and I'm frustrated about losing my post now.
Lets get to the terminology part:
Strand of dna : the instruction set conatained within a single gamete (ignoring MrDNA of course), a haploid's entire dna, half a diploids dna, one side of the double helix (n1).
Double Helix : the pattern created when 2 strands of dna join to form a diploid (or dihaploid) dna set, (n2).
A coding strand: well given all dna is coding, whether its a gene (a segment of a signle strand), an entire strand ie the contents of a haploid nucleus, or part of the garbage dna (segment of the strand again). Just part of the program that when run, creates the plant.
Chromosome : that part of the single dna strand that chooses to stay together, when the strand tears itself apart during both meiosis and mitosis ready to recombine again later. Containing both genes and garbage dna.
Ploidity: the number of stands contained within the nucleus, ie n1 haploid, n2 diploid, n3 triploid, n4 tetraploid etc.
Template strand: A term I haven't come across or used before, therefore I'll go with whatever definition you'd like to give it.
Dihaploid plant: a plant that is created from haploid cells but now contains not the original 1 strand of dna but 2 identical strands (ignoring the odd rewriting mistake here and there created in the process of mitosis. Homozygous for all gene sequences (including garbage dna)
Diploid plant: a plant created by the natural combining of 2 gametes, which will have differences in the 2 strands of dna due to the meiosis process (although I accept that these diffs in some cases will be infinitesimally small).
Mitosis: the process by which a plant grows, the parent cell creating 2 daughter cells and mixes up which strand each chromosome is situated upon.
Meiosis : the process by which gametes are produced (ovums and pollen). The parent cell creates 4 cells each of which only have 1 strand of dna, and that dna is altered in 4 ways (rearrangements, increases in content, decreases in content and mistakes in the copying.)
I'm happy to fine tune any of the above for the purposes of future discussions, as if we're gonna be talking about this in depth, I think it is important we agree on the usage of the terms.
Hey Mofeta, its really nice to be able to discuss this stuff again, so many who used to hold these conversations have now gone awol from this site. And ignoring the politics side of things, just the mechanical difficulties I just ran into makes it so frustrating at times. Rewriting long technical posts is really annoying.
Mofeta : "What I meant is that the doubling of the haploid cells avoids any reshuffling of the genome of the haploid." yes I do agree that the adding of poison (providing you get the dosage just right for the Haploid plant) will merely create a DiHaploid and not a diploid.
Damn I can't remember what I wrote now lol.
The problem as I see it, is that most of the traits in the canna world, aren't necessarily dominant or recessive, but rather co-dominant. And when working with only one side of the double helix, you are automatically excluding the possibility of those traits being present in the DiHaploid. This also ignores those traits that rely on more than one co-dominant gene, of which I believe there are many. Accepting that (to use your original example) the GS Cookies plant, is not a DiHaploid plant, but rather a diploid, then to try to replicate that plant using haploid cells will exclude half of the dna instruction sets necessary to create the plant.
When you say basically correct, if there is anything at all that you disagree with, please just say so straight away, forget about ego issues, I'm not gonna let that get in the way of a decent discussion at all. And it's necessary to get diffs identified instantly in a discussion of this type as letting them slide will only amplify difficulties later on in the discussion.
Sorry man, I just can't remember what I wrote at all and I'm frustrated about losing my post now.
Lets get to the terminology part:
Strand of dna : the instruction set conatained within a single gamete (ignoring MrDNA of course), a haploid's entire dna, half a diploids dna, one side of the double helix (n1).
Double Helix : the pattern created when 2 strands of dna join to form a diploid (or dihaploid) dna set, (n2).
A coding strand: well given all dna is coding, whether its a gene (a segment of a signle strand), an entire strand ie the contents of a haploid nucleus, or part of the garbage dna (segment of the strand again). Just part of the program that when run, creates the plant.
Chromosome : that part of the single dna strand that chooses to stay together, when the strand tears itself apart during both meiosis and mitosis ready to recombine again later. Containing both genes and garbage dna.
Ploidity: the number of stands contained within the nucleus, ie n1 haploid, n2 diploid, n3 triploid, n4 tetraploid etc.
Template strand: A term I haven't come across or used before, therefore I'll go with whatever definition you'd like to give it.
Dihaploid plant: a plant that is created from haploid cells but now contains not the original 1 strand of dna but 2 identical strands (ignoring the odd rewriting mistake here and there created in the process of mitosis. Homozygous for all gene sequences (including garbage dna)
Diploid plant: a plant created by the natural combining of 2 gametes, which will have differences in the 2 strands of dna due to the meiosis process (although I accept that these diffs in some cases will be infinitesimally small).
Mitosis: the process by which a plant grows, the parent cell creating 2 daughter cells and mixes up which strand each chromosome is situated upon.
Meiosis : the process by which gametes are produced (ovums and pollen). The parent cell creates 4 cells each of which only have 1 strand of dna, and that dna is altered in 4 ways (rearrangements, increases in content, decreases in content and mistakes in the copying.)
I'm happy to fine tune any of the above for the purposes of future discussions, as if we're gonna be talking about this in depth, I think it is important we agree on the usage of the terms.
Last edited:
Hi GMT
The dreaded lost post! It is always the long brilliant ones too. These days when I write a long involved post I compose it on a text editor and paste it into ICMag. Or, if I start what I think will be a short post directly in the reply box, and it turns out long, I copy it to my clipboard real quick. Then if I have to log in again I can just paste it in.
I too enjoy discussing these things, nothing crystallizes my own understanding of a subject better than the exposition of it. Many times in trying to explain something to someone I realize that my own grasp of it is lacking/incomplete. This thread is a good example. Thanks for taking the time to explore with me!
I am also quite pleased with your "if there is anything at all that you disagree with, please just say so straight away, forget about ego issues, I'm not gonna let that get in the way of a decent discussion at all." sentiment, very refreshing. I really hate tiptoeing around stuff in order not to offend people/devolve into ego-based argument. I myself LOVE to be corrected, I consider it a favor when someone clears up a misconception in my thinking.
I'm glad you want to leave the DH breeding talk aside for now, it is quite a complex subject. To discuss it properly we need to nail down some fundamentals.
I asked you to explore the terms I mentioned so I could tell where you were at in your study of the mechanisms of inheritance. I suspected that a common misconception was present.
The two strands in a double helix are not inherited separately, one from the M and one from the F, and have nothing to do with ploidy.
In a double helix, both strands come from the same parent, and are complimentary. The individual strands are "reflections" of each other (for lack of a better term), and are not seperate gene sequences.
The chromosome is one long double helix. You get the whole chromosome from one parent. Diploid organisms have two sets of chromosomes, one from each parent. It is the number of sets of chromosomes that determines ploidy.
For example, when you say "The parent cell creates 4 cells each of which only have 1 strand of dna," this is incorrect. The 4 gametes have one set of chromosomes each, and each chromosome in the set is a long double stranded piece of DNA.
The best way I can think of to understand this is to look up the terms coding strand and template strand, and read a number of different definitions of chromosome and ploidy.
I hope I am not being too opaque on this, it is complicated and I have a hard time expressing it with the limited time I have to write here.
Later I will try to find and link to some resources on the web that I feel are particularly illustrative of this concept, it is crucial.
Thanks again for the stimulating discussion!
mofeta
The dreaded lost post! It is always the long brilliant ones too. These days when I write a long involved post I compose it on a text editor and paste it into ICMag. Or, if I start what I think will be a short post directly in the reply box, and it turns out long, I copy it to my clipboard real quick. Then if I have to log in again I can just paste it in.
I too enjoy discussing these things, nothing crystallizes my own understanding of a subject better than the exposition of it. Many times in trying to explain something to someone I realize that my own grasp of it is lacking/incomplete. This thread is a good example. Thanks for taking the time to explore with me!
I am also quite pleased with your "if there is anything at all that you disagree with, please just say so straight away, forget about ego issues, I'm not gonna let that get in the way of a decent discussion at all." sentiment, very refreshing. I really hate tiptoeing around stuff in order not to offend people/devolve into ego-based argument. I myself LOVE to be corrected, I consider it a favor when someone clears up a misconception in my thinking.
I'm glad you want to leave the DH breeding talk aside for now, it is quite a complex subject. To discuss it properly we need to nail down some fundamentals.
I asked you to explore the terms I mentioned so I could tell where you were at in your study of the mechanisms of inheritance. I suspected that a common misconception was present.
The two strands in a double helix are not inherited separately, one from the M and one from the F, and have nothing to do with ploidy.
In a double helix, both strands come from the same parent, and are complimentary. The individual strands are "reflections" of each other (for lack of a better term), and are not seperate gene sequences.
The chromosome is one long double helix. You get the whole chromosome from one parent. Diploid organisms have two sets of chromosomes, one from each parent. It is the number of sets of chromosomes that determines ploidy.
For example, when you say "The parent cell creates 4 cells each of which only have 1 strand of dna," this is incorrect. The 4 gametes have one set of chromosomes each, and each chromosome in the set is a long double stranded piece of DNA.
The best way I can think of to understand this is to look up the terms coding strand and template strand, and read a number of different definitions of chromosome and ploidy.
I hope I am not being too opaque on this, it is complicated and I have a hard time expressing it with the limited time I have to write here.
Later I will try to find and link to some resources on the web that I feel are particularly illustrative of this concept, it is crucial.
Thanks again for the stimulating discussion!
mofeta
Lol yeah, I learned my lesson, typed this up separately in word then copied and pasted. And I agree that if I’m missing something, having that missing knowledge added to the collection is doing me a favour, not putting my nose out of joint. So on with the discussion J
I’m actually happy to continue the DH breeding conversation, but I agree that we need to come to some sort of agreement on the basics first, in order to look into anything more deeply. I’m actually not too bad on this stuff, but neither am I a geneticist, so there are going to be massive gaps here and there. One case in point follows:
Now when you say that “The two strands in a double helix are not inherited separately, one from the M and one from the F, and have nothing to do with ploidy. In a double helix, both strands come from the same parent, and are complimentary. The individual strands are "reflections" of each other (for lack of a better term), and are not seperate gene sequences.”,
Here we go, straight into the deep end hey. You identified a gap right there, yes I was working with an incomplete knowledge of double helix structures within a nucleus. I had made the assumption that you stated. And so now need to update my glossary of terms in the previous post, but don’t want to ruin the continuity of the thread, so will leave my error for all to see instead. It’s easy to hear of the term double helix, and since we all know that we get half our dna from one parent and half from the other, and that the dna is contained within the double helix structure, that one half (strand) is paternal and the other maternal. I had also considered each individual chromosome to exist on one side of the helix or the other, and that they matched up in this double helix structure. I stand educated. I spent the day reading up on this topic believing you to be wrong initially. Though I’d like to clarify one point, the last one, one strand is a sequence of separate genes, and also contains garbage dna sequences. Only one strand is used to create the rna that is then responsible for the protein production. As you said, the other merely being a reflection of that strand. It is this that lead me to my former erroneous conclusion.
You said “The chromosome is one long double helix.” At one point in the cells life this is true, however at most times of the cells life, things are less organised than this.
When you said “For example, when you say "The parent cell creates 4 cells each of which only have 1 strand of dna," this is incorrect. The 4 gametes haveone set of chromosomes each, and each chromosome in the set is a long double stranded piece of DNA.” Yes the use of the term one strand of dna, was incorrect, however the principle basis of what I was getting at is still true. Each gamete only contains half of the initial information, or program for life, (regardless of how it is stored). The objection I was raising is still valid I feel.
I’m actually happy to continue the DH breeding conversation, but I agree that we need to come to some sort of agreement on the basics first, in order to look into anything more deeply. I’m actually not too bad on this stuff, but neither am I a geneticist, so there are going to be massive gaps here and there. One case in point follows:
Now when you say that “The two strands in a double helix are not inherited separately, one from the M and one from the F, and have nothing to do with ploidy. In a double helix, both strands come from the same parent, and are complimentary. The individual strands are "reflections" of each other (for lack of a better term), and are not seperate gene sequences.”,
Here we go, straight into the deep end hey. You identified a gap right there, yes I was working with an incomplete knowledge of double helix structures within a nucleus. I had made the assumption that you stated. And so now need to update my glossary of terms in the previous post, but don’t want to ruin the continuity of the thread, so will leave my error for all to see instead. It’s easy to hear of the term double helix, and since we all know that we get half our dna from one parent and half from the other, and that the dna is contained within the double helix structure, that one half (strand) is paternal and the other maternal. I had also considered each individual chromosome to exist on one side of the helix or the other, and that they matched up in this double helix structure. I stand educated. I spent the day reading up on this topic believing you to be wrong initially. Though I’d like to clarify one point, the last one, one strand is a sequence of separate genes, and also contains garbage dna sequences. Only one strand is used to create the rna that is then responsible for the protein production. As you said, the other merely being a reflection of that strand. It is this that lead me to my former erroneous conclusion.
You said “The chromosome is one long double helix.” At one point in the cells life this is true, however at most times of the cells life, things are less organised than this.
When you said “For example, when you say "The parent cell creates 4 cells each of which only have 1 strand of dna," this is incorrect. The 4 gametes haveone set of chromosomes each, and each chromosome in the set is a long double stranded piece of DNA.” Yes the use of the term one strand of dna, was incorrect, however the principle basis of what I was getting at is still true. Each gamete only contains half of the initial information, or program for life, (regardless of how it is stored). The objection I was raising is still valid I feel.
Good form GMT!
Yeah, this maybe the most common misconception I have run across in discussing molecular genetics with people. If I had a dollar for every time, I could buy a new light!
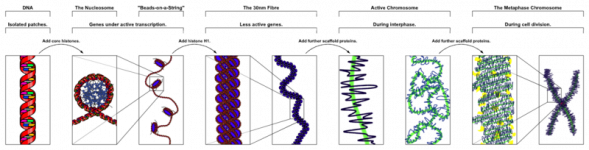
Actually, it is always (except during meiosis) one long double strand. During day to day activity, it is relatively loose. During mitosis and meiosis it condenses and spools up. That's when they have the classic "four arm structure" you see in biology texts. Here is a good illustration I found on the web:

Yeah, there is literally speaking, half of the amount of DNA in a haploid cell, but it is a full instruction set. A diploid cell has two full instruction sets, a triploid three etc. Lots of organisms do perfectly fine with only one set. Drone bees and ants are one example of monoploid/haploid organisms.
In a diploid organism, with two sets, lots of the alleles for a gene are masked by the dominant allele in the other copy of the instructions (chromosome set). So really, there is a difference, but not in the way you are thinking (If I am understanding what your saying above).
Then, if you double a haploid, the result is a truly diploid (two full instruction sets) organism, indistinguishable from a normal diploid, except that the two sets will be perfectly identical (perfectly homozygous), which is basically impossible with traditional breeding.
The term dihaploid you were using earlier refers to diploid individuals derived from tetraploid organisms through culturing gametes or inducer lines or what have you.
I have to go now, I'm pretty busy. Might not be around much for a few days.
Thanks again.
It’s easy to hear of the term double helix, and since we all know that we get half our dna from one parent and half from the other, and that the dna is contained within the double helix structure, that one half (strand) is paternal and the other maternal.
Yeah, this maybe the most common misconception I have run across in discussing molecular genetics with people. If I had a dollar for every time, I could buy a new light!
You said “The chromosome is one long double helix.” At one point in the cells life this is true, however at most times of the cells life, things are less organised than this.
Actually, it is always (except during meiosis) one long double strand. During day to day activity, it is relatively loose. During mitosis and meiosis it condenses and spools up. That's when they have the classic "four arm structure" you see in biology texts. Here is a good illustration I found on the web:
Each gamete only contains half of the initial information, or program for life, (regardless of how it is stored). The objection I was raising is still valid I feel.
Yeah, there is literally speaking, half of the amount of DNA in a haploid cell, but it is a full instruction set. A diploid cell has two full instruction sets, a triploid three etc. Lots of organisms do perfectly fine with only one set. Drone bees and ants are one example of monoploid/haploid organisms.
In a diploid organism, with two sets, lots of the alleles for a gene are masked by the dominant allele in the other copy of the instructions (chromosome set). So really, there is a difference, but not in the way you are thinking (If I am understanding what your saying above).
Then, if you double a haploid, the result is a truly diploid (two full instruction sets) organism, indistinguishable from a normal diploid, except that the two sets will be perfectly identical (perfectly homozygous), which is basically impossible with traditional breeding.
The term dihaploid you were using earlier refers to diploid individuals derived from tetraploid organisms through culturing gametes or inducer lines or what have you.
I have to go now, I'm pretty busy. Might not be around much for a few days.
Thanks again.
Attachments
ah, no, I didn’t make myself clear, sorry. When I said “half the “ that’s what I was referring to. The fact that in the GS Cookies, there are 2 sets of parental information, whereas within the (scientific definition) of your diploid (poisoned haploid) plant (rather than a natural diploid plant) you will only find a doubled single parental contribution. The contents being the same in number of genes, however less in terms of content of genes. In losing half of ( and I accept that half of is inaccurate but it paints the picture better in fewer words) the GSC gene pool, I feel it would become impossible to replicate the traits expressed within the original GSC given as I said earlier, that many traits will not only be co-dominant, but also rely upon multiple genes that are all co-dominant and essential to all be present for the particular trait to manifest. In this sense, no haploid can actually be a full instruction set of the parent, even when doubled, as it is still only one side of an equation. Even though it will still produce a plant. Which is why I raised my objection in the earlier post. And as I said, still feel it’s a valid point that needs addressing, if not here, then in the practical application of what you propose.
I didn’t know about bees or ants, or the origin of the term dihaploid, I merely picked that up from your earlier posts, and misinterpreted how you were using it.
I looked up a couple of you tubes to illustrate what I meant earlier regarding dna
strand creation
http://www.youtube.com/watch?v=z685FFqmrpo
DNA to expressed traits
http://www.youtube.com/watch?v=oYnZiNQlUyo&feature=related
Chromosomes v dna natural state
http://www.youtube.com/watch?v=gbSIBhFwQ4s&feature=relmfu
I didn’t know about bees or ants, or the origin of the term dihaploid, I merely picked that up from your earlier posts, and misinterpreted how you were using it.
I looked up a couple of you tubes to illustrate what I meant earlier regarding dna
strand creation
http://www.youtube.com/watch?v=z685FFqmrpo
DNA to expressed traits
http://www.youtube.com/watch?v=oYnZiNQlUyo&feature=related
Chromosomes v dna natural state
http://www.youtube.com/watch?v=gbSIBhFwQ4s&feature=relmfu
Nunsacred
Active member
I'd like to add some perspective.
Someone said earlier that a trait might be only available if the two parental contributions are both present, which is true, this might happen quite a lot.
So DH populations might entirely lack the desired trait.
^^ This is true.
Think about it, it's more likely to be just 'general healthiness' that gets 'broken' by losing a haplotype, more likely to be a struggle to find a healthy enough phenotypes there.
not so great as a practical breeding tool.
It will help us identify good markers to assist with 'normal' breeding, so it's all good, but I'm just saying ......
...even if we got the best case scenario - pollen giving a good %age of DH seeds from our favourite mother plants -
...it probably isn't useful to a gardener like me...I'd struggle to actually grow the DH plants successfully, from the sounds of things.
Someone said earlier that a trait might be only available if the two parental contributions are both present, which is true, this might happen quite a lot.
So DH populations might entirely lack the desired trait.
^^ This is true.
Think about it, it's more likely to be just 'general healthiness' that gets 'broken' by losing a haplotype, more likely to be a struggle to find a healthy enough phenotypes there.
not so great as a practical breeding tool.
It will help us identify good markers to assist with 'normal' breeding, so it's all good, but I'm just saying ......
...even if we got the best case scenario - pollen giving a good %age of DH seeds from our favourite mother plants -
...it probably isn't useful to a gardener like me...I'd struggle to actually grow the DH plants successfully, from the sounds of things.
wrongwrong
Member
I'm under the impression that it does not matter!, the plant in question can be heterozygous, therefore recombination and linkage is not an issue, so whilst "classical" breeding is all about homologous recombination this isn't. (but this is more to do with Reverse breeding, than what Spurr's initial course was/is.. where he planned to produce dihaploid progeny from DH parental lines).The problem as I see it, is that most of the traits in the canna world, aren't necessarily dominant or recessive, but rather co-dominant. And when working with only one side of the double helix, you are automatically excluding the possibility of those traits being present in the DiHaploid. This also ignores those traits that rely on more than one co-dominant gene, of which I believe there are many. Accepting that (to use your original example) the GS Cookies plant, is not a DiHaploid plant, but rather a diploid, then to try to replicate that plant using haploid cells will exclude half of the dna instruction sets necessary to create the plant.
Surely this sort of application is useful for putting elite's out in mass... and the artificial seed route would not be required.
wrongwrong
Member
Hmmmm, is classical breeding about that, or is it about getting a high proportion to exhibit desired traits? I don't feel they're the same thing at all.
Well if it was about getting a high proportion to show the desired traits you will not get to it faster than methods like this, selfing would be next... it would'nt be in a M/F method producing slow lines after lines taking years.. and even using M/F lines you would not be getting the homozygosity that you will here.... odds in your favour and time too... at the end of the day we want elite genotypes in our gardens showing us elite phenotypes.. (on a side note with homozygous lines/elite specimans etc you could find out what exact enviroment has an effect on any such plant, as such you'd know exactly how epigenetics effect the said plant).
Obviously for me it is impractical, I'm just a simple layman using my simple means which include selfing, I'm not getting a lab anyday soon... so I'll keep chasing my own tail... do you not think those that can incorp this type of tech should? I know you like all things "natural" but hey times they are a changing..
Well don't get me wrong, (sorry no pun intended), I'm not for one second suggesting that the Haploid plants wont show ANY desirable traits. I'm just saying that many of the traits being chased, wont be present in them if trying to replicate a diploid mother plant. You may well find many many new desirable traits, and the plants may well be very healthy, and yes, selfing a plant that started out as a haploid, will create a very stable offspring line, though femmed only, and so can never create a true breeding line without going through the supermale/male/male and female route, which would by definition ruin the goals. I know what the market likes at the moment, but what the market wants, and what's good for the future health and sustainability of the lines may not be the same thing. But that is a different concern. My original point raised, was that the process of meiosis throws the objectives into a little spin, and that by essentially throwing away one half of the pieces to the jigsaw, then completing the same jigsaw again becomes impossible. Though you may end up with a very nice little piece of the jigsaw that is much easier to complete time and time again.
Its way out of my reach too, and Mofeta clearly knows his stuff, and far more about this than I do, I just love these threads though, as they take the site beyond the typical "nice plant man" type of responses to read. Which I also post, and enjoy receiving but its nice to learn stuff too, and toss ideas around that makes everyone re-think things, and that's never a bad thing for anyone. As for should those who can, do, well, with a few provisions in place yes, of course they should. Namely that the work is seen in context, and that it's not the only work being done. But with so many pollen chuckers, breeders, and science bods out there all doing their own thing with our beloved plants, I'm sure we have the diversity of plants and methodologies to maintain a healthy population base.
Its way out of my reach too, and Mofeta clearly knows his stuff, and far more about this than I do, I just love these threads though, as they take the site beyond the typical "nice plant man" type of responses to read. Which I also post, and enjoy receiving but its nice to learn stuff too, and toss ideas around that makes everyone re-think things, and that's never a bad thing for anyone. As for should those who can, do, well, with a few provisions in place yes, of course they should. Namely that the work is seen in context, and that it's not the only work being done. But with so many pollen chuckers, breeders, and science bods out there all doing their own thing with our beloved plants, I'm sure we have the diversity of plants and methodologies to maintain a healthy population base.
wrongwrong
Member
I fully agree, I get bored with most things/threads on here, I need/want more interesting debate etc... though I need putting in place like we all do, I wish there was more top down feeding or such.. but then no one learns thru spoon feeding just more direction needed I guess.. spk soon.Its way out of my reach too, and Mofeta clearly knows his stuff, and far more about this than I do, I just love these threads though, as they take the site beyond the typical "nice plant man" type of responses to read. Which I also post, and enjoy receiving but its nice to learn stuff too, and toss ideas around that makes everyone re-think things, and that's never a bad thing for anyone.
Hi everyone
Boy you guys are bringing this thread to life!
I happened to fall into a large job for someone unexpectedly yesterday, so I won't be able to post much for a while. I have some time to burn right now so I thought I'd jump in. I'll only be able to reply to about the first half of your posts, though.
Remember, I don't intend to do this technique, and I don't recommend it to anyone.
I just made the thread as an intellectual exercise because spurr was interested in discussing it. Also to illustrate to spurr that this was a LOT of work, and had limited context in which it was useful. Especially the centromere mediated technique.
Gamete culture could be done by a serious ameteur at home (with a lot of work), but the centromere stuff is professional lab/expert technician stuff only. This will be (maybe already is) done by big professional outfits like GW. Not by me though. The standard breeding techniques produce results that satisfy me.
That being said, on to discussion.
I totally understand your point, and it is indeed a good question, sorry I didn't make that clear. (As a note, the term polygenic is a useful term to describe traits controlled by multiple genes. Also, instead of "co-dominant" you should say "dependent on" or "influenced by". The term co-dominant has a very specific, precise meaning. It is indeed one of the ways that genes will effect each other, but there are a lot of ways that genes influence each other.)
Here is why I think it doesn't matter. It is the process you brought up a few posts back- meiosis. That is why I wanted to explore DNA and chromosomes, because to understand meiosis properly, you have to have a good picture in your mind of these things.
The process of meiosis produces monoploid haploid gametes, the one set of chromosomes each of the four gametes gets is a mixture of the two sets that the plant producing the gametes got from it's parents. Some of them (maybe very, very few) would happen to have the right combination of alleles to show an excellent Cookie phenotype. The rest are culled. There is more to it, and nunsacred's comment is perfect lead in:
That's a feature, not a bug. The perfect homozygousity of the DH plants exposes deleterious recessive alleles, allowing for culling.
Remember, this technique requires good selection from large numbers.
I cannot think of a reason why a diploid obtained from doubling a haploid would be different than a regular plant, except for the perfect homozygousity.
Think about incrossing plants, like tomatoes. Open pollinated varieties are easy to maintain over many many generations because the are so homozygous. They got that way because the seeds in a tomato fruit are all from pollen from the same plant- selfed. They exclude pollen from other individuals by flower morphology- the female parts never grow far enough out to get past and out of the male parts of the same flower (with most cultivated varieties anyway). To get hybrids you have to remove the seed parent's male parts early, and hand pollinate with pollen from another plant.
How can a plant that only selfs have vigor (like all the open pollinated tomatoes I've ever grown, which is a LOT) you ask? It is because the deleterious allele combos have been weeded out, by virtue of being exposed by selfing.
Tom Hill has made posts here somewhere (and I paraphrase loosely) that if he were to undertake making seed lines from elite cuts, he would treat the cut like and incrosser, selfing and such until he had very homozygous lines, culled of deleterious alleles.
Sure, real high numbers of the first few generations would be pretty weak, but some (again, maybe very very few) would be what you want and you use those. This technique is not preservation, it is cut throat inbreeding- purposeful bottlenecking.
Say you used standard breeding techniques on your favorite line of weed. After years of work you have a F20 line that is killer smoke, vigorous and real homozygous- a pack of seeds would come out very similar ("like clones" is a description I have seen in seed catalogs for some strains).
How is this different from a doubled haploid (excluding the perfection of the homozygousity of the DH)?
Boy, I wish I had more time, because the posts you guys made after the ones I just replied to are quite thoughtful, and really do get into the meat of the matter, you all make good points.
Due to the work that fell into my lap, by the next time I can check in you guys will probably be posting pics of you DH lines!
Boy you guys are bringing this thread to life!
I happened to fall into a large job for someone unexpectedly yesterday, so I won't be able to post much for a while. I have some time to burn right now so I thought I'd jump in. I'll only be able to reply to about the first half of your posts, though.
Remember, I don't intend to do this technique, and I don't recommend it to anyone.
I just made the thread as an intellectual exercise because spurr was interested in discussing it. Also to illustrate to spurr that this was a LOT of work, and had limited context in which it was useful. Especially the centromere mediated technique.
Gamete culture could be done by a serious ameteur at home (with a lot of work), but the centromere stuff is professional lab/expert technician stuff only. This will be (maybe already is) done by big professional outfits like GW. Not by me though. The standard breeding techniques produce results that satisfy me.
That being said, on to discussion.
I feel it would become impossible to replicate the traits expressed within the original GSC given as I said earlier, that many traits will not only be co-dominant, but also rely upon multiple genes that are all co-dominant and essential to all be present for the particular trait to manifest. In this sense, no haploid can actually be a full instruction set of the parent, even when doubled
I totally understand your point, and it is indeed a good question, sorry I didn't make that clear. (As a note, the term polygenic is a useful term to describe traits controlled by multiple genes. Also, instead of "co-dominant" you should say "dependent on" or "influenced by". The term co-dominant has a very specific, precise meaning. It is indeed one of the ways that genes will effect each other, but there are a lot of ways that genes influence each other.)
Here is why I think it doesn't matter. It is the process you brought up a few posts back- meiosis. That is why I wanted to explore DNA and chromosomes, because to understand meiosis properly, you have to have a good picture in your mind of these things.
The process of meiosis produces monoploid haploid gametes, the one set of chromosomes each of the four gametes gets is a mixture of the two sets that the plant producing the gametes got from it's parents. Some of them (maybe very, very few) would happen to have the right combination of alleles to show an excellent Cookie phenotype. The rest are culled. There is more to it, and nunsacred's comment is perfect lead in:
Someone said earlier that a trait might be only available if the two parental contributions are both present, which is true, this might happen quite a lot.
So DH populations might entirely lack the desired trait.
^^ This is true.
Think about it, it's more likely to be just 'general healthiness' that gets 'broken' by losing a haplotype, more likely to be a struggle to find a healthy enough phenotypes there.
That's a feature, not a bug. The perfect homozygousity of the DH plants exposes deleterious recessive alleles, allowing for culling.
Remember, this technique requires good selection from large numbers.
I cannot think of a reason why a diploid obtained from doubling a haploid would be different than a regular plant, except for the perfect homozygousity.
Think about incrossing plants, like tomatoes. Open pollinated varieties are easy to maintain over many many generations because the are so homozygous. They got that way because the seeds in a tomato fruit are all from pollen from the same plant- selfed. They exclude pollen from other individuals by flower morphology- the female parts never grow far enough out to get past and out of the male parts of the same flower (with most cultivated varieties anyway). To get hybrids you have to remove the seed parent's male parts early, and hand pollinate with pollen from another plant.
How can a plant that only selfs have vigor (like all the open pollinated tomatoes I've ever grown, which is a LOT) you ask? It is because the deleterious allele combos have been weeded out, by virtue of being exposed by selfing.
Tom Hill has made posts here somewhere (and I paraphrase loosely) that if he were to undertake making seed lines from elite cuts, he would treat the cut like and incrosser, selfing and such until he had very homozygous lines, culled of deleterious alleles.
Sure, real high numbers of the first few generations would be pretty weak, but some (again, maybe very very few) would be what you want and you use those. This technique is not preservation, it is cut throat inbreeding- purposeful bottlenecking.
Say you used standard breeding techniques on your favorite line of weed. After years of work you have a F20 line that is killer smoke, vigorous and real homozygous- a pack of seeds would come out very similar ("like clones" is a description I have seen in seed catalogs for some strains).
How is this different from a doubled haploid (excluding the perfection of the homozygousity of the DH)?
Boy, I wish I had more time, because the posts you guys made after the ones I just replied to are quite thoughtful, and really do get into the meat of the matter, you all make good points.
Due to the work that fell into my lap, by the next time I can check in you guys will probably be posting pics of you DH lines!
Hey mofeta, well because while many of the genes that we want will be at diff loci, some wont be, and then these become an either or situation unless we can move their loci in some controllable way. In the example you give, in each generation is the possibility to have both ( and in many cases as we have agreed, many) genes present without that either or situation arising. Therefore after many generations, there will be many fine examples of the original within numbers. For the DH tech to work, it would be necessary to marry one set, with the opposing set, which with numbers, genetic testing equip, and the ability to prevent recombination at meiosis, can, it now appears, be done. The resulting offspring being a perfect copy every time. Therefore the DH tech, with the right knowledge and access, is the far superior one. Though I don't know how much the beans would cost after all that.
It is my understanding that 5% (there's that 5% again ) of DH plants may be recombined into a reverse breeding scenario. That doubling haploids doesn't quite get us home (boy all the big names are here ) But yes, this would render all tissue culture and clones fairly obsolete imo.
) of DH plants may be recombined into a reverse breeding scenario. That doubling haploids doesn't quite get us home (boy all the big names are here ) But yes, this would render all tissue culture and clones fairly obsolete imo.
